IF5=7.8 |骆驼蓬属(Peganum)染色体水平基因组组装与群体遗传进化分析
“中国科学院西北高原生物所在《Molecular Ecology Resources》期刊发表最新研究论文,文章通过组装染色体水平物种基因组,群体重测序等分析,首次揭开了骆驼蓬属物种的起源、分化历史及适应性进化机制,并提出创新性的“混合-隔离-再混合 ”(MIM)循环模型”

01
—
研究背景
骆驼蓬属是中亚干旱区的典型沙生属植物类群,包含3个形态各异的物种:骆驼蓬(Peganum harmala L.)、多裂骆驼蓬(P. multisectum (Maxim.) Bobrov)和骆驼蒿(P. nigellastrum Bunge),这些物种的分布区域在河西走廊存在重叠,并在该地区发现了骆驼蓬属的杂交中间体(IHEs),杂交中间体(IHEs):是指在杂交物种形成过程中,介于稳定物种与短暂杂交个体之间的过渡性群体,这些群体在遗传结构上介于亲本之间,可能表现出适应性性状(如抗逆性增强),但未达到独立成种的生殖隔离标准。

为了探究骆驼蓬属物种生物多样性的形成机制,中国科学院西北高原生物所在《Molecular Ecology Resources》期刊发表最新研究论文,文章通过组装染色体水平物种(多裂骆驼蓬)基因组,重测序,群体遗传进化,GWAS等分析,首次揭开了骆驼蓬属物种的起源、分化历史及适应性进化机制,并提出创新性的“混合-隔离-再混合 ”(MIM)循环模型
02
—
研究结果
1. 基因组组装
文章首先采用三种不同的测序技术(ONT、illumina、Hi-C)对青藏高原特有物种多裂骆驼蓬进行了染色体水平的基因组组装,共有98.64%的测序序列定位到了12条染色体上,最终组装得到的基因组大小为314.37Mb,Contig N50为26.99Mb。文章选取了同源序列比对,从头预测和转录组预测三种方式对基因组进行注释,共预测到了22,342个蛋白质编码基因。
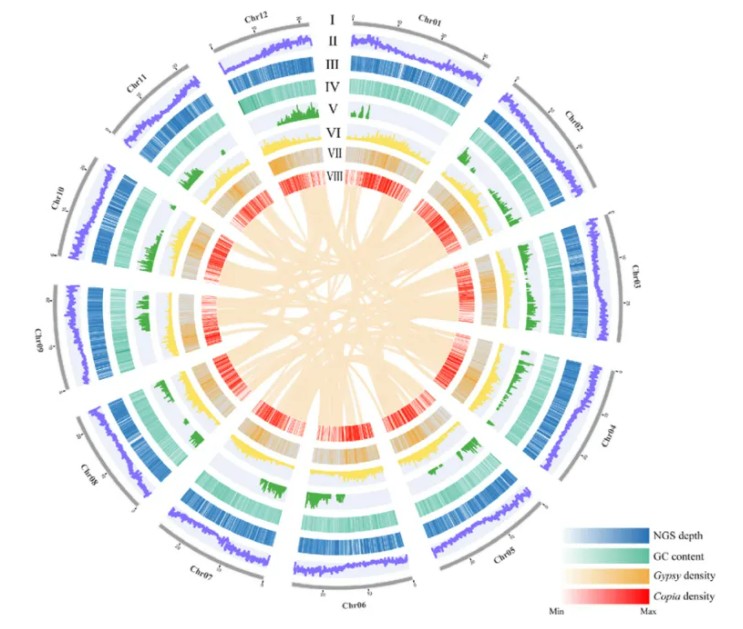
多裂骆驼蓬基因组共线性圈图
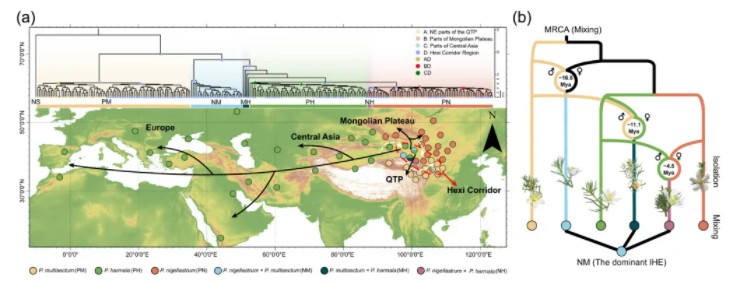
2. 群体遗传进化分析随后作者对中亚干旱区及周边地区采样得到的264个骆驼蓬属物种进行重测序分析,这些样本包括多裂骆驼蓬(89个),骆驼蓬(68个),骆驼蒿(70个),以及这三种物种的杂交中间体(37个),下图为这些样本的采样地分布。
264个多裂骆驼蓬样本采样地分布
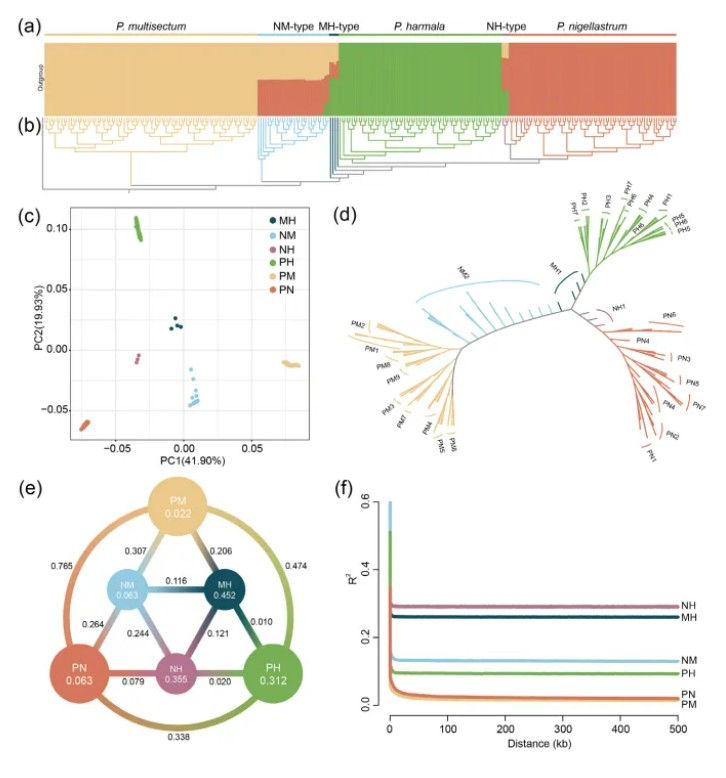
基于264个样本中筛选到的8百多万个SNPs位点,文章进行了群体遗传进化分析,系统发育树构建结果表明,三类杂交中间体物种均聚类于假定的亲本分枝之间。这与主成分分析结果是一致的,这些杂交中间体分布在三个亲本物种中间,并且具有明显的遗传背景差异(c图)。群体结构分析结果计算得到最佳分群数量为K=3,说明这些骆驼蓬属植物的遗传背景是相互独立的。E图展示了各个群体内和群体间计算的核苷酸多样性值(π值)和群体分化系数(FST值),从结果中能看出,相比亲本群体,杂交中间体表现出更高的杂合度,和更大的遗传多样性(π值),以及更广泛的全基因组LD(f图),然而其中的NM群体表现出与其假定亲本PN和PM之间存在显著的遗传分化(F>0.25),提示NM可能处于物种形成的中期,而MH和NH则处于物种形成的早期阶段。
3. 种群历史动态分析
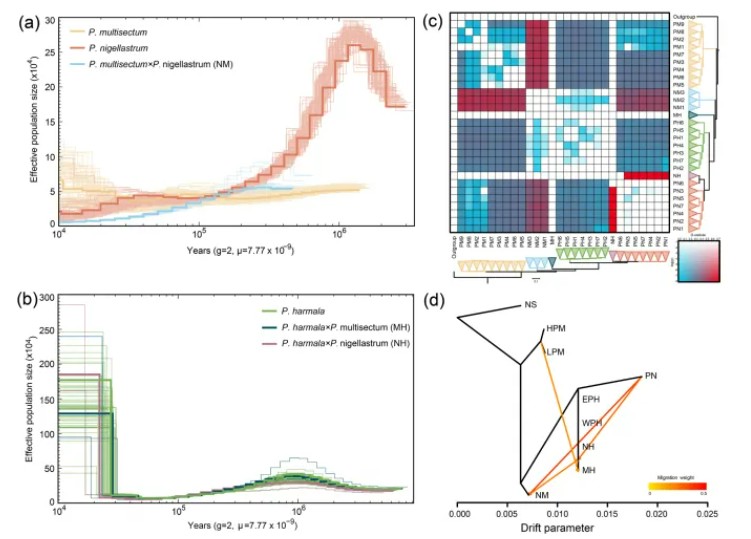
随后作者进行了种群历史动态分析:PSMC分析显示,骆驼蓬属(Peganum)在约150万年前快速扩张,可能与青藏高原的隆升事件有关,同时,青藏高原的隆升导致地理隔离,多裂骆驼蓬(PM)从祖先种中分化出来。TreeMix(d图)分析结果表明,在各种骆驼蓬属植物的融合区域(如河西走廊)等地存在显著基因流动,例如NM和NH从骆驼蒿(PN)处获得基因流,MH从多裂骆驼蓬(PM)处获得基因流,进一步证明河西走廊为基因流的枢纽地带,部分杂交中间体如NM、NH通过基因渐渗获得亲本适应性特征,D统计结果显示NM与 骆驼蒿与多裂骆驼蓬组合时呈现高D值,表明显著渐渗。
4. 分化基因组岛
为了研究进化过程对基因组分化模式的影响,作者对基因组中的分化岛进行了鉴定,基因组分化岛就是基因组中分化程度显著高于中性背景的分化区域,这些区域与物种分化,适应和生殖隔离等进化过程密切相关。为了进一步分析分化岛的成因,作者计算了分化岛和非分化岛中核苷酸差异度(Dxy),核苷酸多样性(π),tajima’D和重组率等参数,结果表明在三种亲本骆驼蓬属植物中(a图),分化岛区域的绝对分化指数,核苷酸多样性显著低于背景区域,且重组率升高,提示自然选择,尤其是长期连锁选择是亲本物种分化的主导因素。而杂交中间体(IHEs)的D统计检验结果中出现了显著的基因渗入信号,表明种间基因流与连锁选择共同塑造了杂交中间体基因组的分化岛。
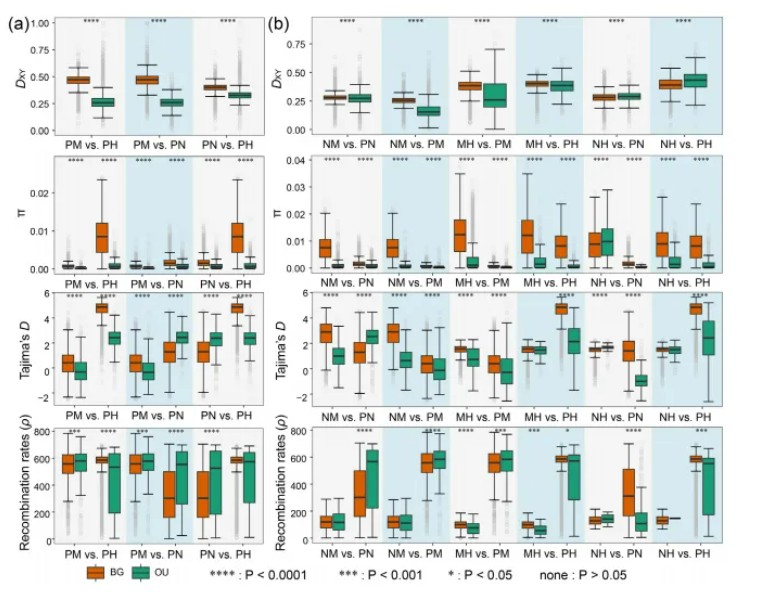
基因组分化岛
5.适应性性状的遗传与表达调控
为了探究骆驼蓬高海拔适应性的遗传基础,文章整合了全基因组选择清除、基因组分化岛和转录组数据,以识别驱动物种分化的适应性基因。基因组选择清除发现多裂骆驼蓬的部分DNA修复基因(如POLD1)受到了强烈的选择作用(图b),并且表现出高度的分化(图c)。随后作者关注了杂交后代对于亲本适应性性状的遗传与继承,GWAS分析定位到骆驼蓬蜡质合成基因FAR的非同义突变(G到C,图e),该突变与蜡质层增厚有关,蜡质层具有减少水分蒸腾、提高抗旱性和减轻紫外线伤害的能力,从而有助于物种适应高海拔地区(图f和图g,是蜡质层合成过程中的相关过程、通路和基因),研究发现该突变在NM群体中部分得以保留并表现出适应性优势,以上结果体现了杂交中间体对于亲本适应性性状的继承与创新。
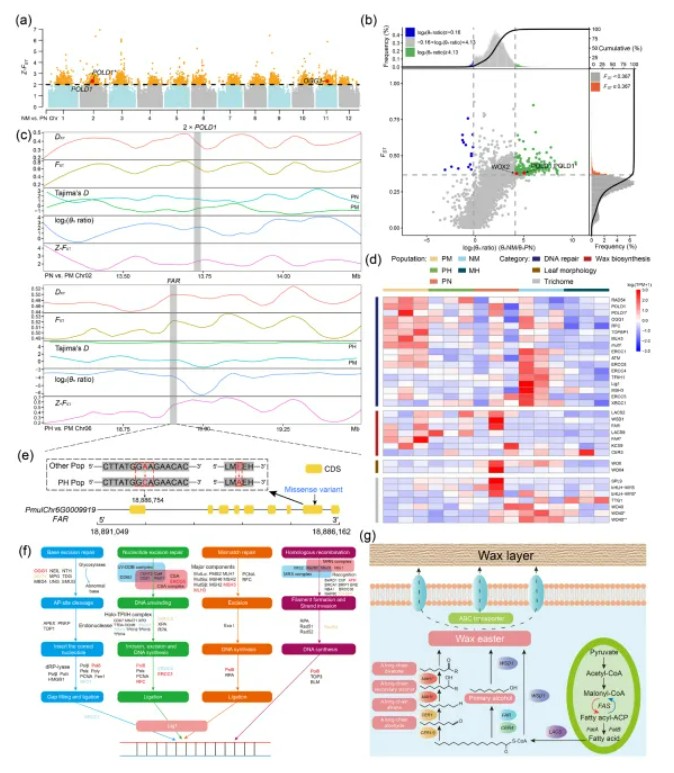
6.基因家族收缩扩张分析
作者选取了包括多裂骆驼蓬在内的多个无患子目的物种以及其他被子植物物种共13种,构建了系统发育关系及分化时间树,并且进行了基因家族收缩扩张分析,图中的饼图展示了每个基因组中收缩和扩张的基因家族所占的比例(绿色表示扩张,红色表示收缩),对多裂骆驼蓬扩张基因家族进行GO和KEGG富集分析,富集结果中发现了与高海拔适应相关的DNA修复以及紫外线调控通路,并且相关性状可能被杂交后代所继承。
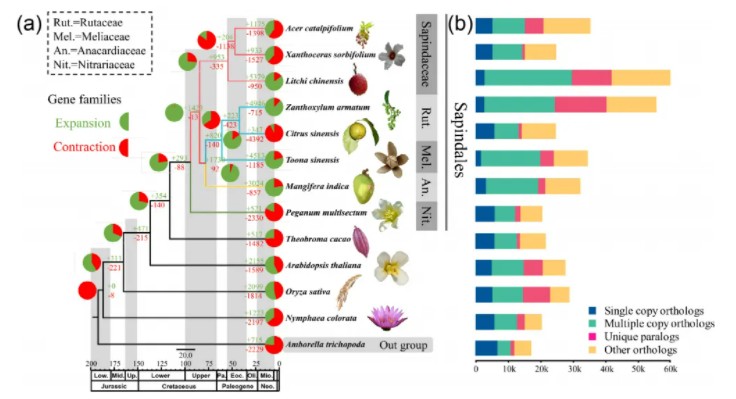
基因家族收缩扩张分析结果
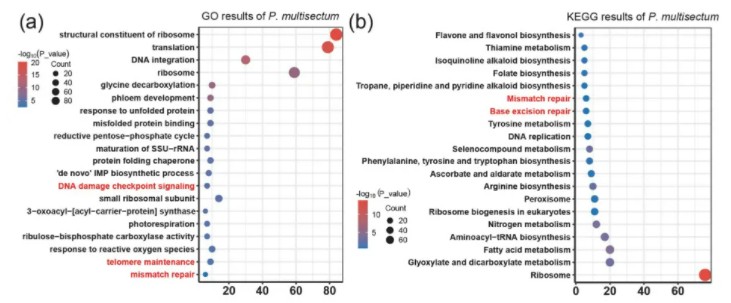
多裂骆驼蓬扩张基因家族富集结果
7.祖先区域重建
为了探究骆驼蓬的起源和演化,作者进行了祖先区域重建分析,分析结果表明骆驼蓬祖先起源于河西走廊地区,约1900万年前分3支扩散至青藏高原、蒙古高原及中亚。研究提出混合-隔离-混合循环(MIM)模型(图b):即物种通过“扩散-地理隔离-再接触杂交”循环演化,在这个过程中IHEs作为“进化种子”整合了亲本优势性状(如NM型兼具耐旱与抗紫外线能力)。b图对文章提出的骆驼蓬属演化轨迹,即混合-隔离-再混合循环(MIM)模型进行了很好的诠释。
- 发表于 2025-05-30 17:09
- 阅读 ( 1466 )
- 分类:文献解读
